
广告
导读
天康生物作为国内第一家猪瘟E2亚单位疫苗的研发、生产和推广单位,我们一直致力于我国猪瘟的深度防控和净化工作。因此,我们一直高度关注各地猪瘟的散发情况,对公开文献报道的临床案例进行定期检索和研究。
本期跟大家翻译分享的是云南省畜牧兽医科学院姚俊老师团队对云南省2015-2021年猪瘟病毒流行病学调查及基因型分析的研究,公开发表于J Vet Sci 2022 Jul;23(4)。
该研究的主要发现:
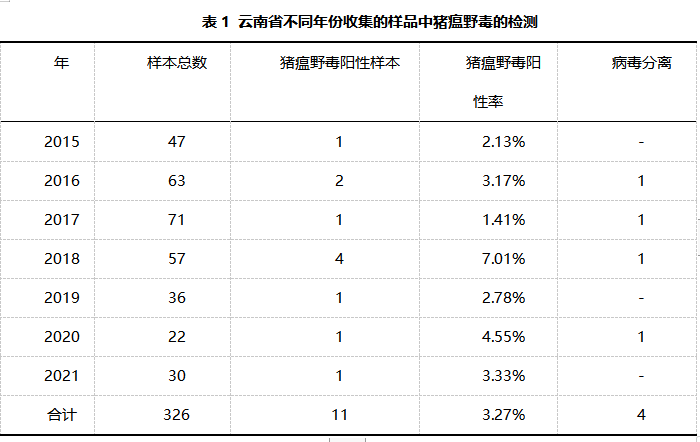
1. 尽管云南省猪群已经接种了猪瘟疫苗,但猪瘟的发病率仍然有3.37%(11/326)。低于山东省(11.1%,188/4866)和中国其他地区,包括河南、河北、黑龙江和辽宁省(6.0%,21/350)。
2. 11株临床分离的猪瘟野毒有2.1c和2.1d两种基因亚型,其中以2.1c基因亚型为主。与公开序列的其他基因亚型2.1c和2.1d的同源性分别为90.0-98.7%和89.6-99.0%。与其他基因2型亚型的序列同源性为85.1-93.0%,包括2.1a、2.1b、2.2和2.3。
3. 11株临床分离的猪瘟野毒与疫苗毒株亲本毒石门毒株(亚型1.1)的同源性较低仅为82.2-83.0%,表明所有猪瘟野毒毒株与疫苗毒株具有不同的亲缘关系。
Epidemiological investigation and phylogenetic analysis of Classical Swine Fever virus in Yunnan province from 2015 to 2021.
云南省2015-2021年猪瘟病毒流行病学调查及基因型分析
全文链接:https://sci-hub.st/10.4142/jvs.22042
摘要
背景:猪瘟是由猪瘟病毒引起的一种高度传染性疾病,对中国养猪业构成严重威胁。
目的:中国许多省份,如山东、河南、河北、黑龙江、辽宁等都报道了猪瘟病毒的流行,而云南省关于猪瘟病毒流行的报道很少。本研究旨在了解云南省猪瘟病毒的流行特征。
方法:本研究于2015年至2021年在云南省不同地区收集了326份组织样本。对326例临床标本进行逆转录聚合酶链反应(RT-PCR)、序列分析、系统进化树分析等病原学检测和分析。
结果:通过RT PCR检测结果表明云南省猪瘟病毒阳性率为3.37%(11/326),低于中国其他地区。对11株猪瘟病毒E2部分序列进行分析,结果表明核苷酸(nt)同源性为80.9 ~ 100.0%,氨基酸(aa)同源性为95.0 ~ 100.0%。系统发育进化树分析新分离株属于2.1c和2.1d基因亚型,以2.1c为主。
结论:2015年至2021,猪瘟病毒在云南省呈散发。该区域发现了基因亚型2.1c和2.1d,但以2.1c为主。
关键词
猪瘟病毒;序列分析;基因亚型2.1c;云南省
简介
猪瘟病毒(猪瘟病毒)属于黄病毒科瘟病毒属,是一种单链正链被膜RNA病毒,基因组长约12.3kb。猪瘟是由猪瘟病毒(猪瘟病毒)引起的一种高度传染性和致死性疾病。该病呈现急性、亚急性、慢性和持续感染型,主要取决于病毒毒力和宿主因素。发生猪瘟会导致高发病率和死亡率,并严重影响养猪业的发展。因此,猪群中出现猪瘟必须向世界动物卫生组织(OIE)报告。猪瘟病毒基因组仅编码一个开放阅读框,经转录和翻译后加工可进一步水解为4个结构蛋白(C、Erns、E1和E2)和7个非结构蛋白(P7、Npro、NS2、NS3、NS4A、SN4B、NS5A和NS5B)。在猪瘟病毒基因组中,具有较高遗传多样性的E2基因被认为是研究猪瘟病毒遗传特征的标准化分子标记。
近年来,全球流行的猪瘟病毒毒株被分为3个基因型(1-3)和11个基因亚型(1.1-1.4、2.1-2.3和3.1-3.4)。猪群中发生猪瘟主要是以2.1基因亚型为主,与其他基因亚型[9]相比,2.1基因亚型表现出更高的遗传变异(2.1a-2.1j)。更重要的是,在许多接种c株疫苗的养猪场监测到了2.1d基因亚型毒株的流行情况。进一步的调查表明,c株疫苗不能完全预防2.1d株。猪瘟在世界不同地区广泛存在,最近在日本就出现了暴发。在中国,尽管猪瘟减毒疫苗(c株)在猪群中广泛应用,但猪瘟疫情仍然存在。为了了解云南省猪瘟疫情的最新情况,2015年至2021年从该地区收集了326份疑似猪瘟组织标本进行调查。对获得的猪瘟病毒新毒株的E2序列进行了遗传特征分析。
材料与方法
尸体剖检:所有猪均严格按照相关标准进行屠宰和解剖。猪被电击瞬间昏迷,失血过多而死。解剖步骤可以归纳为:尸体的外部检查;打开腹腔,检查腹腔脏器;打开胸腔,检查胸前器官。这些步骤由不同农场的兽医执行。
细胞和试剂:猪睾丸细胞(ST)在添加不含BVDV特异性抗体的5%新生牛血清(NBCS)、链霉素(100 μg/mL)和青霉素(100 IU/mL)的培养基(MEM),37℃、5%二氧化碳培养箱中培养。作者实验室保存了石门毒株、c株和抗猪瘟病毒 E2蛋白单克隆抗体(mAb)。
标本的采集和预处理:2015年3月至2021年4月,从云南省不同地区收集了73个疑似感染猪瘟猪场的326个组织(扁桃体和淋巴结)样本(表1)。标本的匀浆在无菌磷酸盐缓冲盐水中进行了三次冷冻和解冻循环。10,000×g在4°C下离心10分钟,收集上清液,保存于−80°C。
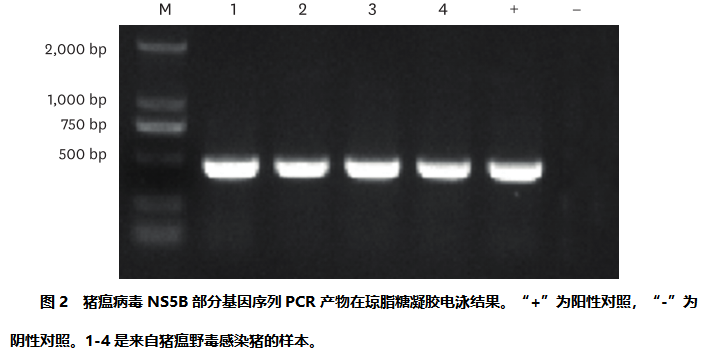
对送样中猪瘟病毒的检测:根据产品说明书,使用商业试剂盒(TaKaRa,日本)从上清液中提取病毒RNA。设计了特异性引物,采用逆转录-聚合酶链式反应(RT-PCR)检测NS5B基因是否存在猪瘟野毒[14]。RT-PCR反应程序:50°C 30 min, 94°C 2 min;35个循环,94°C 30秒,56°C 30秒,72°C 40秒;最后72°C 7分钟。在1%琼脂糖凝胶电泳中,PCR产物的预期DNA条带为449 bp,表明该样品为猪瘟野毒阳性。
猪瘟病毒在ST细胞中的分离与鉴定:将11个具有代表性的猪瘟野毒阳性样本上清液经0.22μm过滤器(Merck Millipore, Cork, Ireland)过滤,接种于单层ST细胞上培养72 h,最后通过RT-PCR和间接免疫荧光法(IFA)验证感染性病毒的存在。猪瘟野毒阳性细胞通过连续传代进一步增殖。在第十次传代时,使用针对猪瘟病毒E2蛋白的单克隆抗体(mAb)通过IFA测定病毒滴度,如前所述[15]。在这项研究中,FITC标记的E2蛋白购自韩国MEDIAN Diagnostics公司。FITC标记的山羊抗小鼠Ig G(二级抗体)购自北京宝龙免疫科技有限公司。FITC标记的羊抗猪二级抗体购自SBA公司。
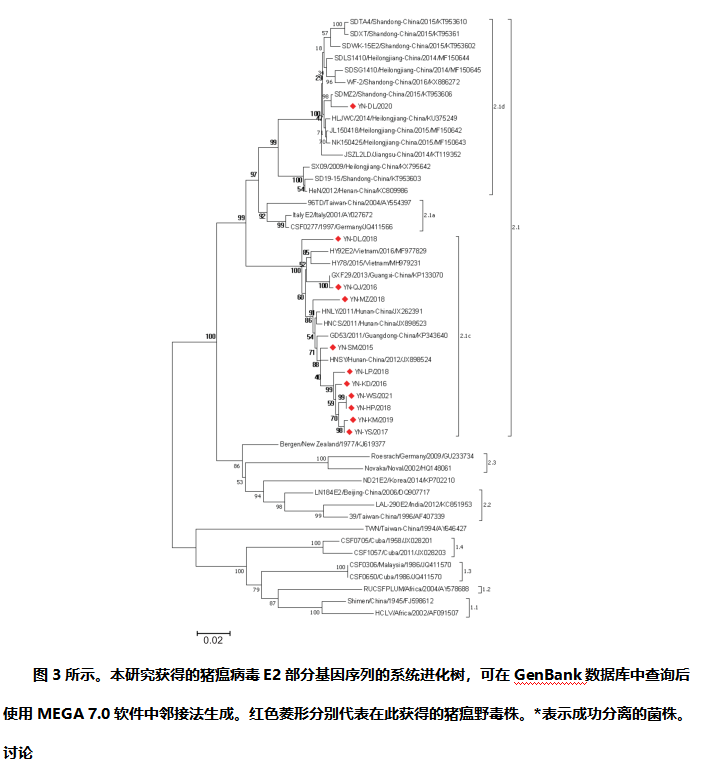
部分E2基因序列分析:用E2- f: 5-GTAAATATGTGTGTGTTAGACCAGA-3 '和E2-r: 5-GTGTGGGTAATTGAGTTCCCTATCA-3 '引物,对11株猪瘟野毒 E2部分基因进行RT-PCR扩增。扩增后,将纯化的PCR产物克隆到pMD19-T载体(TakaRa, Japan)。经酶切鉴定,阳性质粒含目标序列,送测序。将分离的猪瘟野毒毒株的核苷酸序列保存在GenBank数据库中(表2)。用DNA Star Version 7.0软件对分离的11株猪瘟病毒与GenBank数据库中其他地区/国家的参考毒株的E2核苷酸序列进行比对,分析其遗传特征。采用邻接法(Neighbor-Join,NJ),在MEGA 7.0软件中进行1000次构建E2基因序列的系统进化树。
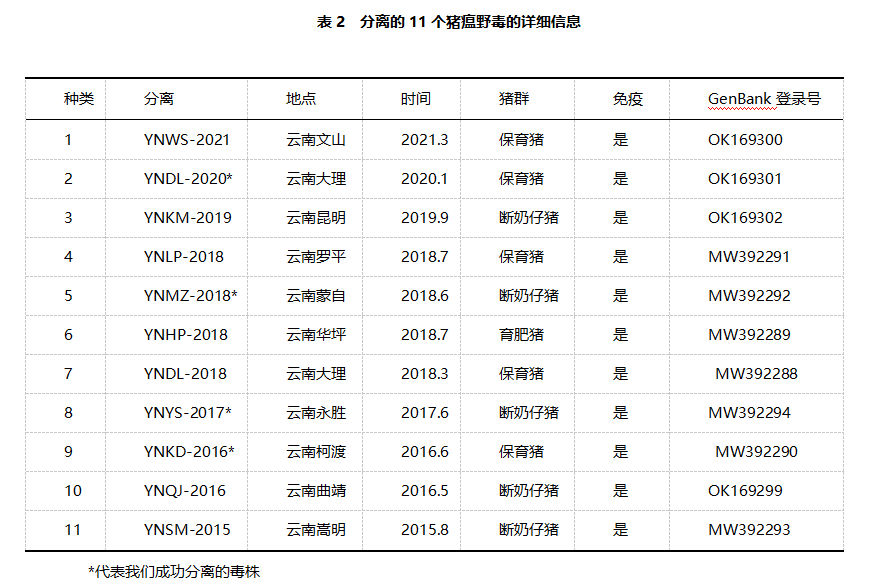
*代表我们成功分离的毒株
试验批准:本研究所有与动物有关的实验均经云南畜牧兽医学院和云南热带亚热带动物病毒病实验室批准。
结果
病理变化:本研究中,从尸检中观察到明显的猪瘟野毒感染病理病变,如皮肤瘀斑(图1A和B),扁桃体溃疡(图1C),淋巴结瘀点(图1D),肾脏瘀点出血(图1E),脾脏梗死(图1F),胃和肠坏死(图1G和H)和回盲瓣按钮状溃疡(图1I)。
猪瘟野毒检测:用PR-PCR检测猪瘟病毒的感染情况。表1列出了2015年至2021年的详细检测结果。共抽取样本326份,猪瘟阳性11份,占3.27%。并且重成功分离到4株猪瘟病毒。图2为猪瘟病毒株NS5B部分基因序列的聚合酶链式反应产物的琼脂糖凝胶电泳图,可观察到大小为449bp的阳性条带。
部分E2基因的系统进化树:本研究对11株猪瘟野毒病毒的E2部分基因序列进行扩增和测序,并将其提交到GenBank数据库,分析云南省猪瘟病毒的遗传特征。表2列出了收集到的11株猪瘟野毒分离株的详细信息。此外,构建了基于E2基因核苷酸序列的邻居连接系统进化树。如图3所示,只有1株猪瘟野毒 (YN-DL/2020)属于2.1d亚基因型。其余10个(YNWS-2021、YNKM-2019、YNLP-2018、YNMZ-2018、YNHP-2018、YNDL-2018、YNYS-2017、YNKD-2016、YNQJ-2016、YNSM-2015)均为2.1c亚基因型。
猪瘟野毒分离株的增殖特性:将猪瘟野毒阳性株的上清接种单层ST细胞,观察其增殖特性。最终成功分离出4株猪瘟野毒 (YNDL-2020、YNKD-2016、YNMZ-2018和YNYS-2017)。利用针对猪瘟野毒 E2蛋白的小鼠单克隆抗体在IFA实验中观察到猪瘟野毒特异性荧光信号(图4)。此外,经15次传代后获得的细胞适应性病毒的感染滴度分别为106.75TCID50/mL、106.375TCID50/mL、107.875TCID50/mL和105.75 TCID50/mL。
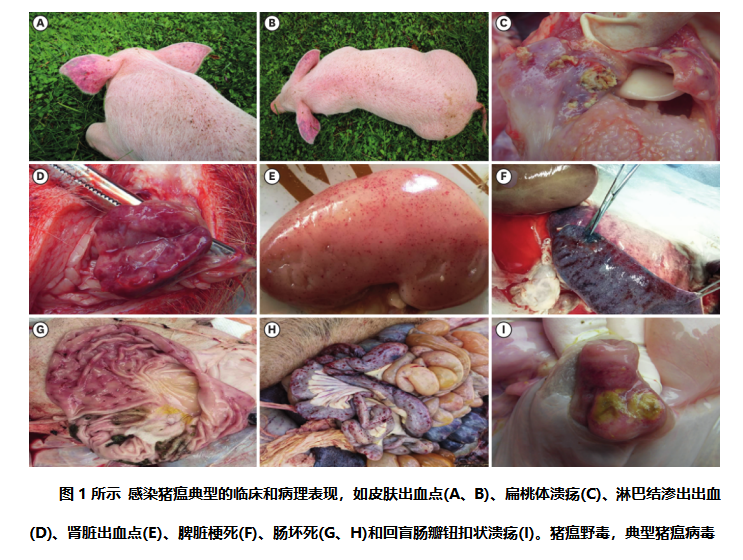
猪瘟野毒感染会导致猪的严重临床症状,包括高热、呕吐、便秘或腹泻,以及皮肤发紫或出血点。确实也在尸检中观察到了明显的病理损害(图1)。在本研究中,只有11个样本经RT-PCR(图2,表1)证实为猪瘟野毒阳性,平均感染率为3.37%(11/326),低于山东省(11.1%,188/4866)[16]和中国其他地区,包括河南、河北、黑龙江和辽宁省(6.0%,21/350)。这些结果表明,尽管云南省猪瘟流行率较低,但这种病原体在中国是普遍存在。这种现象是可以解释的。首先,不同省份的疫情防控政策不尽相同;云南省的阳性率相对较低可能与较好的预防制度和疫苗接种有关。更重要的是,云南是一个多山的省份,森林面积辽阔,为云南省提供了良好的天然屏障,这有利于疫情的预防。云南省的许多农场都位于交通不便的地方,农场分布稀少。
中国流行的猪瘟野毒毒株主要是基因2型,基因2型有2.1、2.2和2.3三个基因亚型。2.1基因亚型比2.2和2.3的流行程度都高,可进一步分为2.1a、2.1b、2.1c和2.1d。特别是2.1基因亚型在中国不同地区的流行呈现出较高的多样性。例如,在山东省主要以 2.1d基因亚型为主,而广东省流行的是2.1c基因亚型。本研究检测了云南省猪瘟病毒的遗传特征,存在2.1c和2.1d两种基因亚型,其中以2.1c基因亚型为主(图3、表2),与广东省情况相似。
扩增出的猪瘟病毒E2基因序列全长为1343bp,编码435个氨基酸。进一步的遗传分析表明,11个毒株在核苷酸和氨基酸水平上的同源性分别为89.0-100.0%和95.0-100.0%。此外,将获得的11株猪瘟野毒与石门株(1.1株)、96TD株(2.1a株)、HNLY株(2.1c株)、SDMZ2株(2.1d株)、LAL-290E株(2.2株)和Novaka株(2.3株)的E2基因序列进行了比较。结果表明,11株猪瘟野毒与基因亚型2.1c和2.1d的同源性分别为90.0-98.7%和89.6-99.0%。此外,它们与其他基因亚型的序列同源性为85.1-93.0%,包括2.1a、2.1b、2.2和2.3。这些新毒株与亚型1.1(石门)的同源性较低仅为82.2-83.0%,表明所有猪瘟野毒毒株与疫苗毒株具有不同的亲缘关系。
对云南省分离的猪瘟病毒株的增殖特性进行了研究。分离到四株猪瘟野毒,并被IFA试验(图4)特异性识别,证明收集的猪瘟野毒可以在单层ST细胞中很好地增殖。有趣的是,YNMZ-2018毒株的病毒滴度是其他毒株的十倍。SA序列比较表明,YNMZ-2018株在E2蛋白中存在一系列独特的氨基酸替换,如49位T到I的变化。此外,在该新毒株中还鉴定出K71R、D97G、K174N、N192S和T197S。然而,这些氨基酸替换是否会影响猪瘟野毒毒株的体外增殖特性还有待进一步研究。
从历史上看,由于中国猪群广泛免疫了有效的猪瘟疫苗,这种传染病似乎在中国的一些地区已经被根除。此外,由于其他传染病的发生或流行,如猪繁殖与呼吸综合征、猪圆环病毒病和猪流行性腹泻,猪瘟本身的预防一直被忽视。在目前的研究中,尽管云南省猪群已经接种了猪瘟疫苗,但猪瘟的发病率仍然有3.37%(11/218)。这表明,这种传染病不容忽视,需要采取相应的预防措施。进一步的遗传分析表明,所有分离的猪瘟病毒都属于2.1c和2.1d亚型,其中云南省主要流行2.1c基因亚型。





