广告
导读:发酵豆粕以其资源丰富、营养较平衡和具有较高的性价比而受到人们的青睐。发酵豆粕是以优质豆粕为主要原料,接种微生物,通过微生物的发酵最大限度地降解豆粕中的抗营养因子,并产生益生菌、寡肽、谷氨酸、乳酸、维生素及未知生长因子等活性物质,具有提高适口性、改善营养物质消化吸收、促进生长、减少腹泻的功效,成为目前鱼粉最好的替代品。本期特邀上海海洋大学博士生导师冷向军教授以“发酵豆粕在水产饲料中的应用研究”为题,介绍了发酵豆粕的营养特性、质量标准,在水产饲料中的应用现状、存在的问题以及对未来的展望,希望对发酵豆粕的优化生产和科学应用提供参考。全文已在《饲料工业》2022年第4期刊出。
鱼粉资源的紧缺和鱼粉价格的上涨,使得替代鱼粉蛋白源的研究一直是水产饲料行业关注的焦点和热点。在这方面,发酵豆粕以其资源丰富、营养较平衡和具较高的性价比而受到青睐。近年来,有关发酵豆粕在水产饲料中的研究和应用不断增多。文章拟就此作一综述,为发酵豆粕在水产饲料中的合理应用提供参考。
1、发酵豆粕的生产和营养特点
豆粕固态发酵是指在人工控制条件下,在豆粕中接种特定的微生物,通过其生长繁殖和新陈代谢作用,去除豆粕中的抗营养因子,提高多肽、小肽及游离氨基酸含量,同时增加益生菌及其代谢产物的过程。常用的发酵微生物有细菌和真菌两大类。细菌类主要有芽孢杆菌、乳酸菌;真菌类主要有酵母菌和霉菌(根霉、毛霉、木霉、曲霉)。根据发酵过程中是否需氧,可分为厌氧发酵(如乳酸菌)和需氧发酵(如芽孢杆菌)。菌种经过逐级扩大培养后和水按比例混合后接种到豆粕中进行厌氧或需氧发酵。
随着固态发酵技术的发展,多菌种联合多级发酵、微生物和酶混合发酵(菌酶发酵)等技术已应用于发酵豆粕,以提升发酵豆粕的营养价值,更好地满足水产饲料的需求。豆粕发酵后,其营养价值的提升主要体现在以下方面。
1.1 减少抗营养因子含量
抗营养因子是指饲料中含有的一些对养分的消化、吸收和利用不利,并使动物产生不良生理反应且影响动物健康和生长的物质。豆粕中含有多种抗营养因子,主要有植酸、胰蛋白酶抑制剂、抗原蛋白、非淀粉多糖、胀气因子、脲酶、大豆凝集素、抗维生素因子和单宁等。这些抗营养因子通过不同的生理机制影响水产动物对营养物质的消化吸收。某些抗营养因子会和饲料中的营养物质发生络合作用或者抑制某些消化酶的活力,影响营养物质的消化吸收,如植酸、胰蛋白酶抑制剂等;某些抗营养因子则会使养殖动物产生抗原反应并对相关器官造成损伤,如抗原蛋白等。水产动物对抗营养因子具有一定程度的耐受性,当饲料中的抗营养因子超过该品种的耐受度后,就会对其生长、免疫、肠道健康等产生负面影响。
豆粕经过微生物发酵后,最明显的变化就是其中的抗营养因子含量显著降低。微生物在繁殖和生长过程中,会利用大量的非蛋白类抗营养因子作为其营养物质,如非淀粉多糖、低聚糖、植酸等;另一方面,微生物产生的消化酶会降解蛋白类抗营养因子,如胰蛋白酶抑制剂、脲酶、大豆凝集素、大豆抗原蛋白等。以曲霉(Aspergillus usamil)发酵豆粕,可将豆粕中的植酸全部降解(Hirabayashi等,1998)。以乳酸菌(Lactic acid bacteria)发酵豆粕后,脲酶抑制剂和棉籽糖含量降低了80.64%和73.5%(Refstie等,2005)。采用枯草芽孢杆菌:乳酸菌:酵母菌联合发酵豆粕,大豆球蛋白含量从224 mg/kg降为107 mg/kg,中性洗涤纤维、酸性洗涤纤维、黄曲霉毒素和玉米赤霉烯酮含量也显著下降(郭萌萌等,2022)。石慧等(2011)采用枯草芽孢杆菌好氧性前发酵+酵母菌和乳酸菌厌氧性后发酵的方式,得到了抗原蛋白几乎完全降解的发酵豆粕。李莹等(2019)采集了具有代表性的10 种商业发酵豆粕产品及其未发酵豆粕,比较了其主要营养成分和抗营养因子含量,表1汇总了其主要结果。
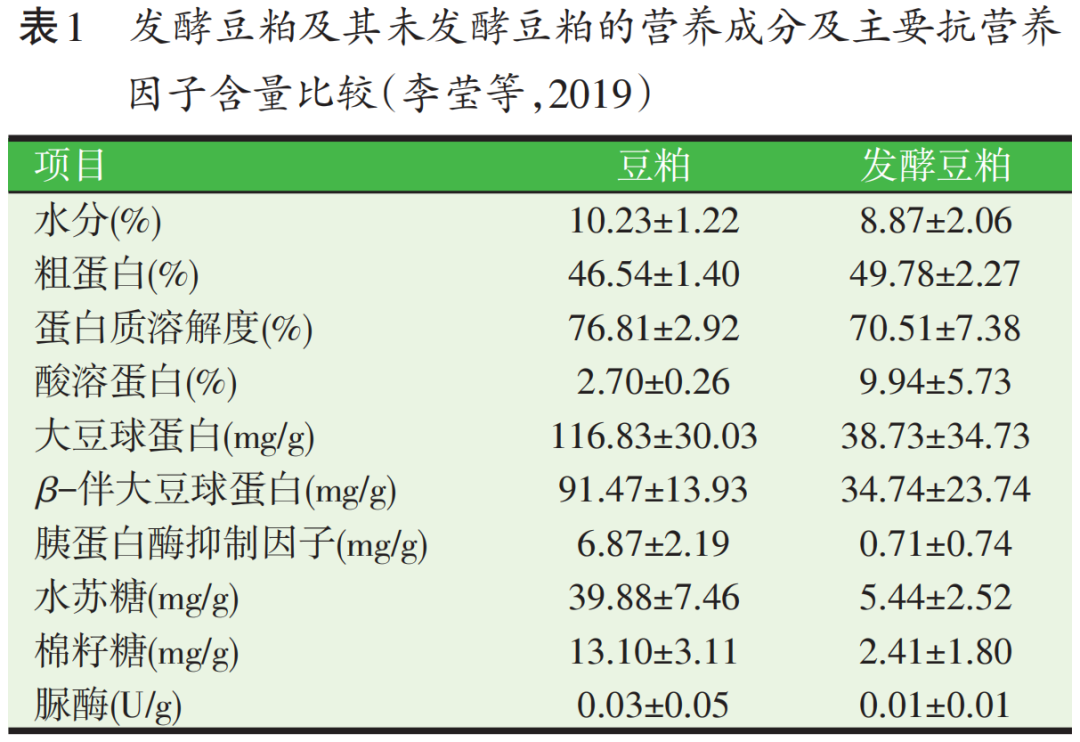
从表1中可见,经过发酵后,发酵豆粕中的主要抗营养因子含量均有了显著降低,其中胰蛋白酶抑制因子、水苏糖和棉籽糖含量的变异程度小,说明这些抗营养因子中的绝大部分已被降解或破坏;而抗原蛋白含量的变异程度较大,表明不同发酵豆粕的发酵工艺、发酵菌种可能存在较大差异,导致抗原蛋白的降解程度不一。酸溶蛋白的增加反映了大分子蛋白的降解;而蛋白质溶解度的下降则表明发酵豆粕生产过程中的烘干环节增加了蛋白质的变性程度。
1.2 提高营养物质和活性成分含量
在发酵过程中,微生物充分利用豆粕中的非蛋白氮和抗营养因子作为生长和繁殖的营养物质来源,合成营养价值更高的菌体蛋白,使得发酵豆粕的粗蛋白质含量增加,相应地,必需氨基酸的组成结构也更为合理。此外,在发酵过程中,微生物的代谢作用消耗了部分有机物料,特别是碳水化合物,出现了蛋白质的“浓缩效应”,使蛋白质含量相对提高。在对10种商业发酵豆粕产品的调查中,李莹等(2019)发现,发酵豆粕的粗蛋白含量(49.78±2.27)%较未发酵豆粕(46.54±1.40)%显著增加(见表1),发酵豆粕中必需氨基酸(苏氨酸、缬氨酸、蛋氨酸、异亮氨酸、亮氨酸、苯丙氨酸、赖氨酸)的平均含量升高2.45%,非必需氨基酸的平均含量升高5.03%。
在发酵过程中,部分大分子蛋白质被降解,产生大小不等的多肽、小肽和游离氨基酸,不仅易于消化吸收,某些成分还具有促生长、促摄食、抗氧化和提高免疫力的作用。豆粕经产朊假丝酵母菌(Candida utilis)发酵后,粗蛋白质含量增加12.4%,游离氨基酸含量增加21倍(陈京华,2006)。以干酪乳杆菌(Lactobacillus casei)发酵豆粕后,酸溶蛋白含量达8.66%,小肽和游离氨基酸含量较发酵前提高510%和280%(吴慧,2016)。小肽和风味氨基酸的增加,使得发酵豆粕风味醇香,具有更好的适口性。李慧娟等(2014)以枯草芽孢杆菌(Bacillus subtilis J3)和植物乳杆菌(Lactobacillus plantarum JNX)发酵豆粕,发酵产物中小肽含量达10.64%,发酵豆粕提取液中的蛋白类产物的分子质量均小于10 ku,其抗氧化活性可达65.76%。以芽孢杆菌固态发酵豆粕48 h后,其总抗氧化活性与多肽含量由207.2 U/g和21.0 mg/g提高到了1 091.1 U/g和347.7 mg/g(徐聪聪,2013)。
微生物的发酵过程,通常伴随着有机酸的产生,如乳酸、丁酸、乙酸、柠檬酸等,这些有机酸随饲料被水产动物摄食后,可在消化道内发挥酸化剂的功能,如促进消化酶的分泌,改善肠道组织结构,维护肠道微生态平衡等。以植物乳杆菌(JUN-DY-6)对豆粕进行固态发酵,可产生6种有机酸,以乳酸、乙酸、丙酸和柠檬酸为主(毛银等,2017)。以乳酸菌固态发酵豆粕,在发酵前期(0~24 h),pH从6.63降为4.34,酸度从0.6%增加到4.19%,之后,pH和酸度的变化则比较缓慢(魏炳栋等,2014)。豆粕经干酪乳杆菌(Lactobacillus casei)发酵后,乳酸含量可达4.64%,总酸含量较发酵前提高了510%(吴慧,2016)。发酵豆粕中的酸度(总酸)和pH可在一定程度上反映出发酵的程度。
大豆异黄酮具有抗菌、抗氧化等生理功能,但豆粕中的大豆异黄酮主要以糖苷形式存在,利用率较低,通过微生物发酵,可将其转化为苷元,显著提高其生理活性。研究表明,微生物发酵可以显著增强大豆异黄酮的抗菌能力和抗氧化能力(杨国峰等,2005;姚明兰等,2003)。李一圣等(2010)以不同菌种发酵豆粕,试验不同程度地降低了3种大豆异黄酮苷类成分(大豆苷、黄豆黄苷、染料目苷)的含量,增加了其苷元类成分的含量。以枯草芽孢杆菌(ATCC6633)发酵豆粕,其苷元含量占大豆异黄酮的比例达到64.56%(杨守凤等,2013)。
豆粕中的磷主要以植酸磷形式存在,发酵过程中,微生物可降解植酸,转化为动物能吸收利用的有效磷。此外,微生物发酵还可以产生酶、维生素和一些未知营养素,这些活性成分也可在一定程度上提高饲料利用率、促进水产动物生长。
2、发酵豆粕标准
目前,我国已制订有行业标准《饲料原料 发酵豆粕》(NY/T 2218—2012),适用于以大豆为主要原料(≥95%),以麸皮、玉米皮等为辅助原料,使用农业农村部《饲添加剂品种目录》中批准使用的微生物菌种进行固态发酵,并经干燥制成的蛋白质原料产品。该标准规定了发酵豆粕的感官性状、技术指标和卫生指标(见表2)。
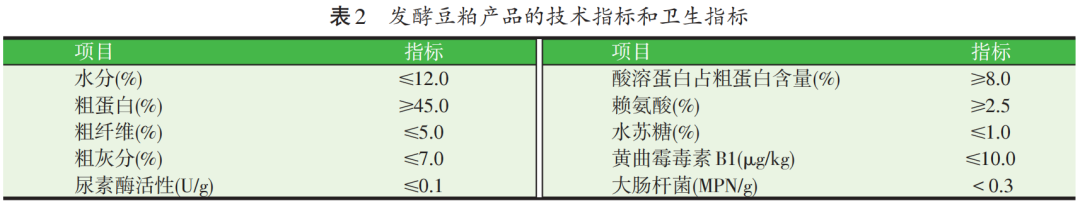
上述指标中,粗蛋白、粗纤维、粗灰分和赖氨酸含量主要由原料豆粕的营养特性决定,尿素酶活性、酸溶蛋白和水苏糖含量则反映发酵程度和抗营养因子降解程度,其中酸溶蛋白含量反映大分子蛋白被分解的情况,生产中多用该指标评价小肽含量,其测定通常是采用三氯乙酸沉淀法,其上清液中的可溶性蛋白含量即为酸溶蛋白含量。黄曲霉毒素和大肠杆菌则反映发酵过程中对安全卫生的控制程度。
3、发酵豆粕在水产饲料中的应用
目前,发酵豆粕在水产饲料中的应用,主要是用作鱼粉的替代蛋白源。肉食性鱼类饲料和虾蟹饲料中鱼粉的用量较高,故发酵豆粕替代鱼粉的研究主要集中在这两大类饲料方面。
3.1 发酵豆粕替代鱼粉的研究
3.1.1 肉食性鱼类
3.1.1.1 大口黑鲈
大口黑鲈是近年发展最为迅速的水产养殖品种之一,2020年的养殖产量已达61.9万吨。大口黑鲈为典型的肉食性鱼类,对鱼粉的要求高,其商业饲料中的鱼粉用量通常为35%~55%。何明等(2020)采用套算法,测定了大口黑鲈对豆粕和发酵豆粕的表观营养物质消化率,在按30%混入基础饲料的条件下,豆粕的干物质、粗蛋白、磷和总氨基酸表观消化率分别为74.71%、76.56%、23.24%和87.34%,发酵豆粕为81.97%、84.0%、46.94%、92.25%,均较豆粕显著提高(P<0.05)。在鱼粉含量为500 g/kg 的饲料中,以芽孢杆菌发酵豆粕替代200 g/kg鱼粉(40%鱼粉),对大口黑鲈(初重19.83 g)的增重和饲料系数没有产生显著影响,但降低了肠绒毛完整性、肠绒毛和杯状细胞的数量;当发酵豆粕替代20%、30%鱼粉时,还提高了增重和特定生长率,其原因主要在于采食量的提高(田鑫鑫等,2021;钟国防等,2021)。同样,在鱼粉含量为500 g/kg的大口黑鲈饲料中,菌酶协同发酵豆粕替代鱼粉比例≤20%时,不会对鱼体生长和饲料利用产生不利影响,替代过高比例鱼粉可对血清生化、免疫和抗氧化指标、肝脏组织形态产生明显的不良影响(陈晓瑛等,2021)。He等(2020a)在鱼粉含量为350 g/kg的饲料中,以多菌种联合发酵豆粕成功替代了105 g/kg鱼粉(30%鱼粉),对大口黑鲈(初重4.43 g)的生长、饲料利用和肠道组织学没有产生显著影响。Yang等(2022)在不补充氨基酸和牛磺酸的条件下,在含鱼粉350 g/kg的大口黑鲈(初重21.2 g)饲料中以乳酸菌发酵豆粕成功替代了100 g/kg鱼粉。
3.1.1.2 鲑鳟鱼类
虹鳟是养殖最为广泛的冷水鱼类,有关发酵豆粕替代鱼粉的研究,主要集中在虹鳟方面。Barnes等(2015)在鱼粉含量为400 g/kg的饲料中,以不同比例发酵豆粕等蛋白替代鱼粉,结果发现,对两个品系的虹鳟而言,含350 g/kg发酵豆粕(鱼粉含量150 g/kg)的饲料均是可接受的,但当发酵豆粕(500 g/kg)全部替代鱼粉后,增重率显著降低,饲料系数显著升高。同样,在鱼粉含量为400 g/kg的饲料中,300 g/kg的发酵豆粕可以成功地替代同等比例鱼粉,尽管饲料的粗蛋白水平有所降低(Barnes等,2012)。Choi等(2020)采用两种发酵程度不同的发酵豆粕,成功地将鱼粉用量从250 g/kg降低到150 g/kg,对虹鳟生长和饲料利用没有产生显著影响。
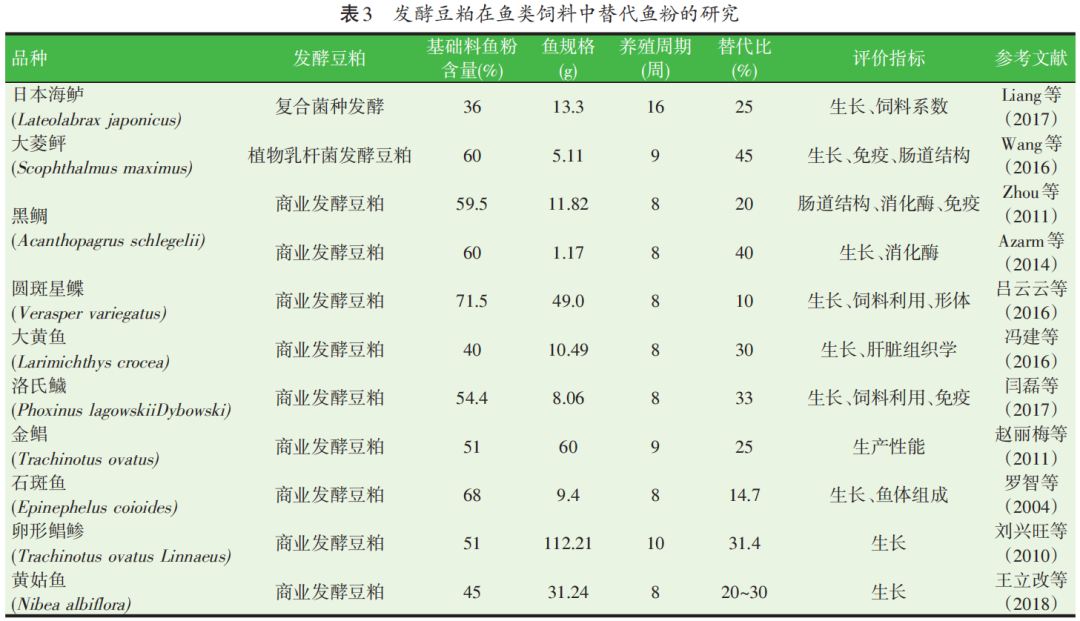
此外,有关发酵豆粕替代鱼粉的报道,也见于黑鲷、大黄鱼、大菱鲆、黄姑鱼、卵形鲳鲹等,详见表3。
3.1.2 虾蟹饲料
和在肉食性鱼类中的研究相比,在虾蟹饲料中应用发酵豆粕替代鱼粉的报道较少。目前,产量最大、应用最广的当属凡纳滨对虾饲料。近年来,凡纳滨对虾饲料中鱼粉用量呈现出逐渐降低的趋势,目前,商业饲料中的鱼粉用量一般为20%左右。Van Nguyen等(2018)、Shiu等(2013)和Yang等(2009)报道,发酵豆粕可成功替代凡纳滨对虾饲料中鱼粉用量的25.4%、61.67%和30%,但在这些研究中,基础饲料中的鱼粉用量均较高,分别为30%、56%和30%。Yao等(2020)设计了鱼粉含量为18%的凡纳滨对虾饲料,发现豆粕和发酵豆粕可分别替代1/6和1/3的鱼粉,发酵豆粕较豆粕能替代更高比例的鱼粉。
在日本沼虾的研究中,根据生长和免疫指标,复合菌种发酵的豆粕可以替代基础饲料(含鱼粉580 g/kg)中25%的鱼粉(Ding等,2015)。Xu等(2020)在鱼粉含量为250 g/kg的饲料中,以两种发酵豆粕分别替代100、150 g/kg鱼粉,对中华绒螯蟹的生长和蛋白质合成、代谢相关基因表达没有产生不利影响。
总体来看,发酵豆粕替代鱼粉的用量,与水产动物的食性、生长阶段、基础饲料配方组成,特别是鱼粉用量等有关。通常,基础配方中的鱼粉用量高,则发酵豆粕可替代的鱼粉量就大,反之亦然。
3.2 发酵豆粕对水产动物生理生化和代谢的影响
目前,有关发酵豆粕对水产动物生理生化和代谢的影响,主要是在替代鱼粉的基础上开展的。
3.2.1 摄食与消化
在一些关于发酵豆粕替代鱼粉的研究中,有发现采食量提高的现象。比如,田鑫鑫等(2021)将芽孢杆菌发酵豆粕以10%、15%、20%、25%的量添加到大口黑鲈饲料中(鱼粉用量由50%降为45%、40%、35%和30%),大口黑鲈采食量较对照组(50%鱼粉组)提高10.7%、10.4%、11.6%、17.8%,但对饲料系数没有显著影响。Yang等(2022)也发现,随发酵豆粕替代鱼粉比例增加,大口黑鲈幼鱼的摄食率呈增加趋势。在大黄鱼的研究中,发酵豆粕替代60%和75%的鱼粉时,幼鱼的摄食率也显著提高(何娇娇等,2018)。然而,在He等(2020)的研究中,并没有报道大口黑鲈采食量增加的现象,这可能与该试验采用等量投喂有关。发酵豆粕替代鱼粉可以提高摄食量,其原因可能与发酵豆粕中的部分小肽和游离氨基酸具有诱食性有关,也可能是与发酵豆粕刺激消化道蠕动促进排空有关。
相比于豆粕,发酵豆粕能够提高水产动物的消化酶活力和消化吸收能力。一方面,经过发酵后,豆粕中的大多数抗营养因子被去除,从而降低了对消化酶的抑制作用;另一方面,发酵豆粕具有的一些活性肽和消化酶对饲料的消化吸收起到了促进作用。Shiu等(2015)在斜带石斑鱼(Epinephelus coioides)的研究表明,替代鱼粉用量的30%后,发酵豆粕组的肠道胰蛋白酶、胰凝乳蛋白酶、淀粉酶和脂肪酶活性均显著高于豆粕组。He等(2020a、2020b)发现,在鱼粉含量为350 g/kg的基础饲料中,以豆粕替代鱼粉用量的30%,显著降低了粗蛋白和干物质消化率,替代60%的鱼粉,显著降低了肠绒毛高度和宽度,但发酵豆粕替代鱼粉的比例达到60%时,才表现出营养物质消化率的下降,并且对肠绒毛高度没有影响(仅宽度有下降)。在圆斑星鲽(Verasper variegatus)的研究中,随着发酵豆粕替代鱼粉比例的升高,胃蛋白酶和肝脂肪酶活力表现为先降低后升高的趋势,而肠淀粉酶活力则先上升后下降,20%发酵豆粕组的肠道淀粉酶活力显著高于对照组,各发酵豆粕组的肠绒毛高度和肠壁厚度均较对照组显著降低(吕云云等,2016)。彭翔等(2012)发现,以20%发酵豆粕替代鱼粉,可提高黑鲷(Acanthopagrus schlegelii)肠道蛋白酶和脂肪酶活性,对肠道结构没有明显损伤。尽管存在一些不一致的报道,但从总体来看,以发酵豆粕替代一定比例的鱼粉,对水产动物的消化功能和营养物质消化率不会产生影响,但过高的替代比例,会损伤消化道结构功能,这可能是鱼粉比例过低后缺少活性成分对肠道的营养作用所致。
3.2.2 免疫与抗病力
豆粕发酵过程中会产生具有免疫调节特性的生物活性肽、寡糖和益生菌等活性物质,这可能是发酵豆粕提高水产动物免疫功能和抗病力的重要物质基础。陈萱等(2005)报道,以含发酵豆粕的饲料饲养异育银鲫(Carassius auratus gibelio),其血清ALT活性显著高于普通豆粕组。彭松等(2017)在凡纳滨对虾上的研究表明,发酵豆粕能提高血淋巴酚氧化酶(PO)和总超氧化物歧化酶(T-SOD)活性。刘韬等(2018)发现,发酵豆粕替代适当比例鱼粉,可提高凡纳滨对虾相关免疫酶活性,上调免疫相关基因表达。然而,当发酵豆粕的用量过高后,则会对水产动物的免疫功能产生不利影响。当发酵豆粕替代鱼粉的比例达到49.5%和66%时,洛氏鱥(Phoxinus lagowskii dybowski)血清中的SOD和溶菌酶(LZM)活性显降低(闫磊等,2017)。
目前,有关发酵豆粕对水产动物抗病力影响的研究较少。Yang等(2022)以225 g/kg发酵豆粕等蛋白替代150 g/kg鱼粉,大口黑鲈感染嗜水气单胞菌(腹腔注射)后的死亡率较对照组显著降低。Lee等(2016)发现,发酵豆粕替代适量鱼粉可提高许氏平鲉感染爱德华氏菌后的存活率。在凡纳滨对虾,发酵豆粕替代适量鱼粉也降低了哈维伊弧菌(Vibrio harveyi)攻毒后的死亡率(Kim等,2017)。在牙鲆的研究中,发酵豆粕替代适量鱼粉增强了非特异性免疫能力(Kader等,2012)。这些研究表明,适当比例的发酵豆粕可提高水产动物的抗病力。
3.2.3 肠道微生物
肠道微生物对鱼类的生长和健康起着重要作用,并受饲料、水环境以及鱼类种类、发育阶段等因素影响。以大口黑鲈为例,在门水平上,肠道微生物主要由梭杆菌门、厚壁菌门、放线菌门和变形菌组成(He等,2020a),在属水平上主要包括鲸杆菌属和支原体属等(He等,2020a;Zhou等,2018)。Yang等(2022)发现,随发酵豆粕替代鱼粉水平升高,大口黑鲈肠道梭杆菌门和鲸杆菌属相对丰度下降,厚壁菌门和支原体属相对丰度则呈上升趋势;当鱼粉水平从350 g/kg降至200 g/kg时,上述趋势基本稳定,而Sobs指数和Chao指数则在250 g/kg鱼粉组最高,反映出物种丰富度的增加。这表明,适量的发酵豆粕可以提高肠道菌群组成的多样性,这可能与发酵豆粕中含有丰富的小肽、益生菌、有机酸以及黄酮类物质有关,它们为不同菌群的生长和繁殖提供的更多的营养环境。在Li等(2020)和Wang等(2019)的研究中,发酵豆粕替代鱼粉,同样提高了大菱鲆和大黄鱼肠道菌群的多样性。Catalan等(2018)认为,发酵豆粕能通过增加肠道微生物群落中乳酸菌的相对丰度,促进大西洋鲑的生长和健康,这种作用类似于益生菌的作用。然而,何娇娇等(2018)发现,发酵豆粕替代鱼粉对大黄鱼肠道微生物的香农多样性指数、辛普森指数和覆盖率没有显著影响。Shao等(2019)在凡纳滨对虾上的研究也表明,发酵豆粕替代鱼粉不会对肠道微生物产生显著影响。这些不同的结果,可能与所用发酵豆粕的种类(发酵菌种、代谢产物)和替代鱼粉的比例,以及水环境等因素有关。
4、问题与展望
4.1 发酵豆粕的营养局限
豆粕经过发酵后,其营养价值有了很大提高,但是,我们也应该看到发酵豆粕营养特性的局限性,这也是限制发酵豆粕在水产饲料中大量使用的重要原因。发酵豆粕由豆粕发酵而来,这决定了发酵豆粕具有豆粕的基本营养特性,比如,蛋氨酸较为缺乏。当发酵豆粕的使用量较高的时候,必须考虑氨基酸的平衡,通过补充外源氨基酸,或通过蛋白质的互补作用而加以实现。此外,作为植物蛋白源,发酵豆粕中仍然缺乏牛磺酸、胆固醇等动物来源的活性成分。当发酵豆粕替代鱼粉的比例较高时,来源于鱼粉的活性成分减少,从而导致消化道的损伤和生产性能的下降。发酵豆粕中含有较多的有机酸,当发酵豆粕的用量过大后,这些有机酸可能会破坏消化道的酸碱平衡。这些都是在发酵豆粕替代鱼粉的研究和应用中需要考虑的问题。
4.2 发酵豆粕的生产工艺和质量标准
目前商业发酵豆粕大都采用单菌种发酵工艺,也有采用多菌种、多级发酵工艺的发酵豆粕产品。不同的生产企业,有不同的生产工艺流程和参数。固态发酵易受环境因素的影响,如温度、湿度、光照等,同时外来杂菌也会在豆粕中繁殖,这些均会影响发酵豆粕的品质。另一方面,固态发酵一般采用批次发酵,不同批次之间的产品品质也会存在差别,导致了产品品质的不稳定。这都需要规范发酵豆粕的生产工艺,以保证产品质量及其稳定性。
在豆粕发酵之前,需要对原料豆粕进行灭菌,如果缺少这个环节,或者灭菌不充分,会导致发酵过程中杂菌的生长和繁殖。在发酵过程中,会产生一些活性物质,这些活性物质中的相当一部分是热敏性的,比如酶、维生素、活的微生物等。在干燥过程中,高温会提高干燥效率,降低生产成本,但对热敏性活性成分的损伤和破坏作用也大,建议尽量采用低温干燥,最大程度地保留活性成分,充分发挥出发酵豆粕的功能性作用。
由于发酵菌种和发酵工艺的不同,其产物的种类和数量不同,降解抗营养因子的程度也有不同。显然,从现有的行业标准来看,并不能完全反映出发酵豆粕的产品特性。今后,有必要分别针对乳酸菌、芽孢杆菌等发酵的豆粕,制订行业标准或团体标准。
4.3 发酵豆粕作用机理的研究
发酵豆粕的营养价值,除了体现在常规营养成分,如蛋白质、氨基酸等方面外,也体现在发酵产物中的活性成分方面。然而,这些活性成分组成复杂,含量也参差不齐,如活性肽、酶、维生素、有机酸、活性微生物、寡糖以及一些未知因子等。如能针对这些成分在发酵产物中的含量,以及在发酵豆粕中的含量进行测定,并研究其单独或协同的作用效果,将有助阐明发酵豆粕的作用机理。在此基础上,通过发酵菌种的筛选和发酵工艺的优化,定向生产富含某种或某几种活性成分的功能性发酵豆粕,将进一步提升发酵豆粕的应用价值。
4.4 由鱼粉替代蛋白源向常规蛋白源的方向发展
目前,关于发酵豆粕的研究应用,绝大多数是用作替代鱼粉的蛋白源。随着发酵豆粕产品的标准化生产和行业对发酵豆粕特性认识的加深,发酵豆粕可望作为一种常规的功能性原料应用于水产饲料中。当然,这需要开展更多的评估工作,以了解不同水产饲料中发酵豆粕的适宜用量,以及不同养殖环境和条件下不同水产动物对发酵豆粕的反应。

最新发布